Details of DPV and References
DPV NO: 155 October 1975
Family: Virgaviridae
Genus: Tobamovirus
Species: Odontoglossum ringspot virus | Acronym: ORSV
Odontoglossum ringspot virus
H. L. Paul Biologische Bundesanstalt für Land- und Forstwirtschaft, Institut für Landwirtschaftliche Virusforschung, Braunschweig, Germany
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
Described by Jensen & Gold (1951), and Paul et al. (1965).
A virus with rod-shaped particles c. 300 x 18 nm. It was first isolated from Odontoglossum grande and can easily be transmitted mechanically. Infects orchids, and a few plants from other families, but host range has not been studied extensively. The virus occurs in cultivated orchids in different parts of the world. No vector known.
Main Diseases
Causes ringspot symptoms in Odontoglossum grande (Fig. 1), and ‘diamond mottle’ in Cymbidium (Jensen & Gold, 1951; Inouye, 1966).
Geographical Distribution
Reported from cultivated orchids in different parts of the world.
Host Range and Symptomatology
The virus infects systemically Odontoglossum grande, species of Cymbidium, Cattleya, Epidendrum and Zygopetalum, Zinnia elegans (no leaf symptoms) and Nicotiana clevelandii. Local lesions without systemic infection occur in Beta vulgaris cv. cicla, Chenopodium amaranticolor, C. quinoa, Gomphrena globosa, Nicotiana glutinosa, N. tabacum cv. Xanthi-nc and Tetragonia expansa. In N. tabacum cv. Samsun the inoculated leaves were symptomlessly infected; Datura stramonium is not infected (Paul et al., 1965).
-
Diagnostic species
- Nicotiana tabacum
cv. Xanthi-nc. Older inoculated leaves show necrotic local lesions 2-4 mm in diameter after a few days; younger leaves produce more ring-like lesions (Paul et al., 1965; Fig. 2, Fig. 3). Both types of local lesion are smaller than those made by type strain and larger than those made by Corbett’s (1967) isolate TMV-O. - The symptoms produced in orchid are not sufficiently reliable to be used as a diagnostic character.
-
Propagation species
- The virus multiplies well in the inoculated leaves of Nicotiana tabacum cv. Samsun. Systemically invaded N. clevelandii may also be useful (Paul et al., 1965).
-
Assay species
- Chenopodium quinoa
reacts with small chlorotic to necrotic spots within a few days after inoculation; Nicotiana tabacum cv. Xanthi-nc may also be used.
Strains
Two isolates studied in detail showed identical behaviour (Paul et al., 1965). The existence of different strains is not yet demonstrated but may explain reported differences in symptomatology and host range.
Transmission by Vectors
No report.
Transmission through Seed
No information.
Transmission by Dodder
No information.
Serology
Moderately immunogenic; antibody titres of 1/1024 have been obtained. Gel-diffusion and ring-precipitin tests give satisfactory results. Gel-diffusion and intragel absorption tests with several other tobamoviruses show the presence of determinants specific to odontoglossum ringspot virus (C. Wetter, unpublished data; Fig. 9, Fig. 10).
Relationships
The virus can be distinguished from most other tobamoviruses by serology and host range. The differentiation of the virus from other tobamovirus isolates from orchid remains doubtful, because a comparative study is lacking (Corbett, 1967).
Stability in Sap
In sap of Nicotiana tabacum cv. Samsun the virus lost infectivity after 10 min at about 90°C or after dilution to 10-5 to 10-6 (Paul et al., 1965).
Purification
The virus can easily be purified by several methods. Clarification of sap with chloroform-butanol followed by differential centrifugation is recommended (Paul et al., 1965). Yield of virus from sap of inoculated leaves of tobacco cv. Samsun is c. 1 g/l.
Properties of Particles
In sucrose density gradients (Paul et al., 1965) virus preparations produce three main zones (Fig. 4), the upper and broader one containing short particles of various lengths (Fig. 5), the middle one containing preponderantly ‘normal’ particles 300 nm long (Fig. 6), and the lower one containing aggregates. All types of particle are identical serologically. The sedimentation constants (s°20,w) are 212 S for the hypersharp peak of 300 nm particles, and c. 119 S for the broad peak of shorter particles (Fig. 8). The short particles are not infective (Paul et al., 1965).
The isoelectric point is not determined, but measurements of turbidity of virus suspensions showed two maxima at pH 3 and at pH 5-6.5. Possibly the long and the short particles aggregate at different pH values.
A260/A280 (corrected for light-scattering): 0.99 for preparations containing predominantly ‘normal’ particles, 0.63 for those containing the short ones. Positions of maximum and minimum for ‘normal’ particles were 268 and 248 nm, and for short particles 280 and 250 nm.
Particle Structure
Particles are rigid rods, having a normal length of c. 300 nm (Paul et al., 1965; van Regenmortel et al., 1964), and a width of c. 18 nm. There is a central canal, and the particles seem to be very fragile: preparations contain many short particles and ‘discs’ (Fig. 7).
Particle Composition
Nucleic acid: The ‘normal’ particles contain c. 5% RNA; many of the short particles are presumably free of RNA because they contain on average only c. 1.2% RNA. Unfractionated virus contains on average c. 2% RNA (Paul et al., 1965).
Protein: The protein subunit has 157 amino acid residues; their sequence
(Hennig, 1972)
is:
10
AcSer -Tyr-Ser-Ile-Thr-Thr-Pro-Ser-Glx-
Leu-Asx-Tyr-Leu-Ser-Ser-Ala-Trp-Ala-Asx-
20 30
Pro-Lys-Glx-Leu-Ile-Asx-Leu-Cys-Thr-Asx-
Ala-Leu-Gly-Asx-Ser-Phe-Glx-Thr-Glx-Asx-
40 50
Ala-Arg-Thr-Thr-Val-Gln-Gln-Gln-Phe-Ala-
Asp-Val-Trp-Thr-Pro-Ser-Pro-Gln-Leu-Thr-
60 70
Val-Arg-Phe-Pro-Ala-Gly-Ala-Gly-Tyr-Phe-
Arg-Val-Tyr-Arg-Tyr-Asx-Phe-Ile-Leu-Asx-
80 90
Pro-Leu-Ile-Thr-Pro-Leu-Met-Gly-Thr-Phe-
Asp-Thr-Arg-Asn-Arg-Ile-Ile-Glx-Val-Glx-
100 110
Asx-Glx-Pro-Asx-Pro-Thr-Thr-Ala-Glx-Thr-
Leu-Asx-Thr-Thr-Arg-Arg-Val-Asp-Asp-Ala-
120 130
Thr-Val-Ala-Ile-Arg-Ser-Ala-Ile-Asn-Asn-
Leu-Leu-Asn-Glu-Leu-Val-Arg-Gly-Thr-Gly-
140 150
Met-Tyr-Asx-Glx-Ser-Thr-Phe-Glx-Val-Met-
Gly-Trp-Thr-Ser-Ser-Leu-Ser-Thr.
M. Wt of the protein subunit is 17,598 daltons (from sequence); 17,300 daltons (from equilibrium centrifugation, Paul & Buchta, 1971).
Relations with Cells and Tissues
The virus induces the formation of crystalline inclusion bodies a few days after inoculation. The bodies consist of one or more stacked plates, composed of virus particles lying perpendicular to the flat surface of the plates. Rarely there are hexagonal prisms similar to those found with type strain TMV, but thinner and not of the typical shape (Milicic & Stefanac, 1971).
Notes
The virus can be distinguished from all other viruses in orchid, except other tobamoviruses, by its particle morphology and its ability to produce necrotic local lesions in Nicotiana tabacum cv. Xanthi-nc.
Acknowledgements
Figures 2 to 8: Biologischc Bundesanstalt für Land- und Forstwirtschaft, Braunschweig; Figure 1 after Jensen & Gold, 1951; Figs. 9 and 10 courtesy of Dr C. Wetter.
Figures

Odontoglossum grande; leaf with ringspot symptoms.

Nicotiana tabacum cv. Xanthi-nc; old inoculated leaf.

Nicotiana tabacum cv. Xanthi-nc; young inoculated leaf.

Bands produced in a sucrose density gradient: broad upper band contains short particles; two sharp lower bands contain preponderantly particles 300 nm long and longer aggregates respectively.

Virus particles from density gradient, upper band. Bar represents 300 nm.

Virus particles from density gradient, lower band. Bar represents 300 nm.
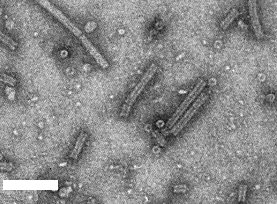
Unfractionated virus particles negatively stained showing central channel and ‘discs’ showing central hole and hexagonal outline. Bar represents 100 nm.

Schlieren diagram produced by moving boundary sedimentation of preparations of odontoglossum ringspot virus top) and type strain TMV (bottom). Note broad peak of short particles in top curve. Sedimentation is from left to right.

Reaction of antiserum to odontoglossum ringspot virus (central wells) with the following viruses: a = odontoglossum ringspot; b = opuntia ringspot; c = sunn-hemp mosaic; d = cucumber green mottle mosaic; e = tobacco mosaic, type strain U1; f = ribgrass mosaic; g = tobacco mosaic strain U2; h = tomato mosaic. (Fig. 9) Agar gel diffusion test; (Fig. 10) intragel absorption test (before the antiserum and the test antigens were applied, the antiserum well was supplied with a mixture of antigens b to h), the remaining homologous reaction shows the antiserum contains a large proportion of antibodies specific to odontoglossum ringspot virus.

Reaction of antiserum to odontoglossum ringspot virus (central wells) with the following viruses: a = odontoglossum ringspot; b = opuntia ringspot; c = sunn-hemp mosaic; d = cucumber green mottle mosaic; e = tobacco mosaic, type strain U1; f = ribgrass mosaic; g = tobacco mosaic strain U2; h = tomato mosaic. (Fig. 9) Agar gel diffusion test; (Fig. 10) intragel absorption test (before the antiserum and the test antigens were applied, the antiserum well was supplied with a mixture of antigens b to h), the remaining homologous reaction shows the antiserum contains a large proportion of antibodies specific to odontoglossum ringspot virus.
References list for DPV: Odontoglossum ringspot virus (155)
- Corbett, Phytopathology: 57: 164, 1967.
- Hennig, in Atlas of Prot. Sequence and Structure 5: D-287, ed.M. O. Dayhoff, Washington, National Biomedical Research Foundation, 1972.
- Inouye, Ber. Öhara Inst. landw. Biol. 13: 149, 1966.
- Jensen & Gold, Phytopathology 41: 648, 1951.
- Milicic & Stefanac, Acta bot. croat. 30: 33, 1971.
- Paul & Buchta, J. gen. Virol. 11: 11, 1971.
- Paul, Wetter, Wittmann & Brandes, Z. VererbLehre. 97: 186, 1965.
- van Regenmortel, Hahn & Fowle, S. Afr. J. agric. Sci. 7: 159, 1964.