Details of DPV and References
DPV NO: 213 July 1979
Family: Bromoviridae
Genus: Cucumovirus
Species: Cucumber mosaic virus | Acronym: CMV
This is a revised version of DPV 1
Cucumber mosaic virus
R. I. B. Francki Department of Plant Pathology, Waite Agricultural Research Institute, University of Adelaide, South Australia
D. W. Mossop Department of Plant Pathology, Waite Agricultural Research Institute, University of Adelaide, South Australia
T. Hatta Department of Plant Pathology, Waite Agricultural Research Institute, University of Adelaide, South Australia
Contents
- Introduction
- Main Diseases
- Geographical Distribution
- Host Range and Symptomatology
- Strains
- Transmission by Vectors
- Transmission through Seed
- Transmission by Grafting
- Transmission by Dodder
- Serology
- Nucleic Acid Hybridization
- Relationships
- Stability in Sap
- Purification
- Properties of Particles
- Particle Structure
- Particle Composition
- Properties of Infective Nucleic Acid
- Molecular Structure
- Genome Properties
- Satellite
- Relations with Cells and Tissues
- Ecology and Control
- Notes
- Acknowledgements
- Figures
- References
Introduction
-
Described by Doolittle (1916) and
Jagger (1916).
Selected synonyms
- Cucumber virus 1 (Rev. appl. Mycol. 6: 501)
- Cucumis virus 1 (Rev. appl. Mycol. 17: 52)
- Marmor cucumeris (Rev. appl. Mycol. 28: 514)
- Spinach blight virus (J. agric. Res. 14: 1)
- Tomato fern leaf virus (Rev. appl. Mycol. 9: 417)
- Cucumis virus 1 (Rev. appl. Mycol. 17: 52)
-
A virus with three functional pieces of single-stranded RNA, packaged in three classes of icosahedral particle about 28 nm in diameter, all sedimenting at the same rate. The virus has a very wide host range and a world-wide distribution; it is transmitted by numerous species of aphid in a non-persistent manner.
Main Diseases
Causes mosaic of cucumber and many other cucurbits, blight of spinach, fern leaf of tomato, mosaic of celery, and mosaics of many other species of dicotyledonous and monocotyledonous crop, ornamental and weed plants including some trees and shrubs. The virus also often causes plant dwarfing and flower breaking (Smith, 1972).
Geographical Distribution
World-wide, especially in temperate regions.
Host Range and Symptomatology
Extremely wide host range; Price (1940) recorded 191 susceptible species in 40 families and Komuro (1973) infected plants in 39 families with the Japanese common strain. Readily transmitted by inoculation with sap, inducing a variety of symptoms depending on virus strain and host cultivar.
-
Diagnostic species
- Chenopodium amaranticolor
and C. quinoa. Chlorotic or necrotic local lesions but no systemic infection. - Cucumis sativus (cucumber). Systemic mosaic
(Fig. 1) and stunting varying in severity.
- Cucurbita moschata (zucchini marrow). Systemic mosaic (Fig. 2) and stunting varying in severity.
- Lycopersicon esculentum (tomato). Mosaic with narrowing of leaf laminae producing characteristic fern leaf symptoms (Fig. 3).
- Nicotiana glutinosa. Symptoms vary with virus strain, ranging from mild (Fig. 4) to severe mosaic (Fig. 6). Some strains induce vein yellowing (Fig. 5) and mosaic.
- Vigna unguiculata (cowpea). Small brown lesions (Fig. 7) in inoculated leaves; some strains infect the plants systemically, inducing mild mosaic.
- Cucurbita moschata (zucchini marrow). Systemic mosaic (Fig. 2) and stunting varying in severity.
-
Propagation species
- Nicotiana glutinosa
and N. tabacum cv. Xanthi-nc are convenient for maintaining cultures. N. tabacum and Cucurbita pepo (pumpkin) are suitable as virus source plants but N. clevelandii appears to be the best source for most strains.Assay species
- Vigna unguiculata
(Crowley, 1954), Chenopodium amaranticolor and C. quinoa are all suitable for local lesion assays.
Strains
Many symptom variants occur, making the virus often difficult to identify from symptoms alone. Strains well characterized by their biological properties include the Y strain which infects cowpea systemically (Price, 1934), the spinach strain which produces severe symptoms on tobacco (Bhargava, 1951) and the M strain originally isolated from Price's (1934) No. 6 yellow strain (Mossop, Francki & Grivell, 1976). In Japan, Komuro (1973) distinguished five strains by their biological properties. Strains characterized physicochemically include the Y strain (Kaper, Diener & Scott, 1965), the Q strain (Francki et al., 1966; Habili & Francki, 1974a) and the S strain (Van Regenmortel, 1967; Van Regenmortel, Hendry & Baltz, 1972). A number of strains have been assigned to two groups, ToRS and DTL, according to their antigenic properties; the Q and S strains both belong to the former group (Devergne & Cardin, 1973, 1975; Devergne, 1975).
Transmission by Vectors
Transmissible in the non-persistent manner by more than 60 species of aphid (Kennedy, Day & Eastop, 1962). Transmission efficiency varies with the aphid species (Simons, 1959) and host plant species (Simons, 1955, 1957). Some virus strains are more readily transmitted than others (Simons, 1957; Normand & Pirone, 1968) and transmissibility by Myzus persicae can be lost altogether (Badami, 1958; Mossop & Francki, 1977). Virus can be acquired by all instars within 5-10 sec; their ability to transmit declines after about 2 min and is usually lost within 2 h (Hoggan, 1933; Watson & Roberts, 1939; Simons, 1955).
Transmission through Seed
Occurs to varying degrees in 19 species (Neergaard, 1977), including some weed species; dissemination and persistence in weed seeds may be important in the epidemiology of the virus (Tomlinson & Carter, 1970).
Transmission by Dodder
Transmitted by at least 10 species of Cuscuta, in which the virus multiplies (Schmelzer, 1957).
Serology
The immunogenicity of the virus is poor but can be enhanced by fixation with formaldehyde (Francki & Habili, 1972). The virus precipitates on exposure to physiological salt solutions and mild heating and therefore serological tests are usually done by agarose double-diffusion tests in buffers of low molarity or in water (Francki et al., 1966; Scott, 1968). Unstabilized virus may give one or two precipitin lines, one nearer the antigen well, caused by more or less intact virus, and a second one nearer the antiserum well caused by degraded virus (Scott, 1968; Devergne & Cardin, 1970).
Relationships
In its physical and chemical properties, the virus has affinities with tomato aspermy virus (Hollings & Stone, 1971) and peanut stunt virus (Mink, 1972), all being members of the cucumovirus group (Matthews, 1979). Cowpea ringspot (Phatak, Diaz-Ruiz & Hull, 1976) and robinia mosaic viruses (Schmelzer, 1971) may also be members. Some members of each of the two serological subgroups of cucumber mosaic virus are related to some strains of tomato aspermy and peanut stunt viruses, but other members of each group show no such relationship (Devergne, 1975; Devergne & Cardin, 1975; Habili & Francki, 1975). There is lack of agreement as to whether cucumber mosaic and tomato aspermy viruses cross-protect against each other (Blencowe & Caldwell, 1949; Hollings, 1955; Holmes, 1956; Graham, 1957; Govier, 1957), but a relationship between the two is indicated by observations that pseudo-recombinants can be made from parts of their genomes (Habili & Francki, 1974b; Mossop & Francki, 1977, 1979). However, nucleic acid hybridization analyses indicate that they have little base sequence homology (Habili & Francki, 1974b; Gonda & Symons, 1978).
Stability in Sap
Relatively unstable in plant extracts, being unable to withstand temperatures in excess of 70°C for 10 min. Infectivity is lost within a few days, and in some instances hours, at room temperature (Smith, 1972; R. I. B. Francki, unpublished data). The virus is inactivated by ribonuclease (Francki, 1968) and infection is strongly inhibited by the presence of leaf extracts from all the plant species tested (Francki, 1964).
Purification
Several methods have been used, most of which are modifications of that devised by Scott (1963). Most virus strains can be purified by one of the following two methods.
1. Modification of Lot et al. (1972). Extract each 100 g leaves with 100 ml 0.5 M sodium citrate (pH 6.5) containing 5 mM EDTA and 0.5% thioglycollic acid, emulsify with 1 vol (w/v) chloroform and centrifuge at 12,000 g for 10 min. To the buffer phase, add polyethylene glycol (M.Wt 6000) to 10% and stir gently for 30-40 min at 0-4°C. Centrifuge at 12,000 g for 10 min and resuspend the sediment in 40-50 ml 5 mM sodium borate buffer, 0.5 mM EDTA, pH 9.0, add Triton X-100 to 2% and stir for 30 min. Clarify by centrifugation at 19,000 g for 15 min and subject the virus to three cycles of differential centrifugation, resuspending the virus pellets in 5 mM borate buffer, 0.5 mM EDTA, pH 9.0. Further purification may be achieved by centrifugation in 5-25% sucrose density gradients (Francki, 1972).
2. Mossop et al. (1976) used the following method for purifying a strain which could not be prepared by Method 1. Extract each 100 g leaves in 300 ml of 0.1 M Na2HPO4 containing 0.1% each of thioglycollic acid and sodium diethyldithiocarbamate (c. pH 8) and centrifuge at 8000 g for 10 min. Add Triton X-100 to the supernatant fluid to a final concentration of 2% (v/v), stir for 15 min at 0-4°C and sediment the virus at 105,000 g for 2 h. Resuspend the sediment in the original extraction medium and subject to differential centrifugation and sucrose density gradient centrifugation as in Method 1.
Virus yields vary according to virus strain and host plant used but over 500 mg virus/kg leaf material has been obtained in some instances.
Properties of Particles
Readily disrupted in the presence of low concentrations of sodium dodecyl sulphate or high concentrations of neutral chloride salts, indicating that the particles are stabilized by RNA-protein interactions (Kaper, 1975).
Sedimentation coefficient: the particles sediment as a single component (Fig. 8) with s20, w (Svedbergs) = 98.6 - 1.04c where c is the virus concentration in mg/ml (Francki et al., 1966).
M. Wt: estimates vary from 5.0 to 6.7 x 106 (Francki et al., 1966; Van Regenmortel, 1967; Dupont et al., 1968; Van Regenmortel et al., 1972). Values based on diffusion coefficients determined by laser light-scattering range from 5.8-6.1 x 106 (D. W. Mossop, unpublished data).
Diffusion coefficient (D20 x 10-7cm2/sec): 1.29-1.35 for four virus strains using laser light-scattering (D. W. Mossop, unpublished data).
Partial specific volume (v'): 0.701 (calculated assuming virus to contain 18% RNA and v' for RNA and protein to be 0.55 and 0.74 respectively).
Electrophoretic mobility: mobility varies with the strain (Lot & Kaper, 1976). That of the S strain is -8 x 10-5 cm2 sec-1 volt-1 at pH 7.0 in 0.1 M buffer; mobility as a function of pH is given by Van Regenmortel (1967).
Extinction coefficient (E (0.1%, 10mm)) at 260 nm: 5.0 (Francki et al., 1966).
A260/A280: about 1.7 (corrected for light-scattering).
Particle Structure
Particles disrupt when mounted unfixed in neutral phosphotungstate for electron microscopy (Francki et al., 1966) and flatten when air-dried prior to shadowing with metal (Scott, 1963). However, particles remain intact and contrast well in uranyl acetate (c. pH 4.5) (Francki et al., 1966). They are isometric, about 28-30 nm in diameter (Fig. 10), and contain 180 protein subunits in pentamer-hexamer clusters with T = 3 surface lattice symmetry (Finch, Klug & Van Regenmortel, 1967). Particles have a hydrated diameter of about 33 nm when measured by laser light-scattering (D. W. Mossop, unpublished data). Neutron small-angle scattering data indicate that the internally located RNA lines the protein shell, leaving a central hole approximately 12 nm in diameter, and penetrates the protein to a depth of about 2 nm (Jacrot, Chauvin & Witz, 1977).
Particle Composition
Nucleic acid: Single-stranded RNA, about 18% of particle weight (Kaper & Re, 1974). G:A:C:U= 24:23:23:30 (Habili & Francki, 1974a). RNA consists of four species of M. Wt c. 1.27 x 106 (RNA-1), 1.13 x 106 (RNA-2), 0.82 x 106 (RNA-3) and 0.35 x 106 (RNA-4), determined by polyacrylamide gel electrophoresis under denaturing conditions described in Fig. 9. M. Wt of the RNA species may differ slightly between strains (D. W. Mossop, unpublished data). RNA-1 and RNA-2 are encapsidated separately and RNA-3 and RNA-4 are in the same particle; thus virus preparations contain particles of at least three different types but with similar morphology and sedimentation properties (Fig. 8). Only the three largest RNA molecules are necessary for infection (Peden & Symons, 1973; Lot et al., 1974); each is a unique molecule except for a common sequence of about 200 nucleotides at each 3'-terminus (Gould & Symons, 1977, 1978). The nucleotide sequence of RNA-4, which contains the coat protein gene (Schwinghamer & Symons, 1977), is present in RNA-3. All four RNA species can be charged at the 3'-end with tyrosine by plant-derived amino acyl tRNA synthetases (Kohl & Hall, 1974) and their 5' ends are ‘capped’ with 7-methyl guanosine (Symons, 1975).
Small, encapsidated, single-stranded RNA molecules of about 1 x 105 daltons have been detected in some isolates of CMV (Kaper, Tousignant & Lot, 1976; Mossop & Francki, 1977; Gould et al., 1978). Their total dependence on CMV for replication and their unique base sequences (Lot, Jonard & Richards, 1977; Diaz-Ruiz & Kaper, 1977; Gould et al., 1978; Mossop & Francki, 1978) indicate that they are ‘satellite’ RNA molecules. One of these, called CARNA 5, appears to be responsible for a lethal necrotic disease of tomato (Kaper & Waterworth, 1977); its complete sequence of 335 nucleotide residues has been determined (Richards et al., 1978).
Protein: Coat protein can be isolated by disrupting virus and precipitating the RNA with 2 M LiCl (Francki et al., 1966). The protein subunit has a M. Wt of c. 24,500 (Van Regenmortel et al., 1972; Habili & Francki, 1974a). The amino acid composition varies slightly between strains of the virus (Linnasalmi & Toiviainen, 1974; Mossop et al., 1976).
Other components: None reported.
Relations with Cells and Tissues
Detection of virus particles by electron microscopy is difficult because of their similar appearance in sections to ribosomes (Tolin, 1977). However, particles have been identified in the cytoplasm, nuclei and vacuoles but not in mitochondria or chloroplasts (Gerola, Bassi & Belli, 1965; Lyons & Allen, 1969; Honda & Matsui, 1968, 1974). Using a technique by which the RNA of ribosomes is digested without affecting that of the virus (Hatta & Francki, 1979), particles, some of which appear hollow, can be seen scattered throughout the cytoplasm of infected cells (Fig. 11). Sometimes the particles aggregate into crystalline arrays, usually in vacuoles (Fig. 12); these aggregates have also been detected by light microscopy (Christie & Edwardson, 1977).
A virus-specific RNA-dependent RNA polymerase has been isolated from infected plants (Clark, Peden & Symons, 1974) and double-stranded replicative forms corresponding to the four viral RNA species have been detected in infected protoplasts (Takanami, Kubo & Imaizumi, 1977) and leaves (Kaper & Diaz-Ruiz, 1977).
Notes
The physical, antigenic and biological properties of the cucumoviruses overlap so as to make identification of many isolates difficult. It is usually considered that tomato aspermy virus can be distinguished from cucumber mosaic virus by its ability to invade systemically Chrysanthemum but not cucumber, and by its ability to cause tomato to set seedless fruits and to induce enations on leaves of Nicotiana spp. (Hollings & Stone, 1971). Peanut stunt virus has been considered to be distinguishable by its ability to invade cowpea, French bean and peanut systemically (Mink, 1972). However, these criteria appear not to be infallible. For example, some virus isolates antigenically very similar to the Q strain of cucumber mosaic virus infect cowpea systemically whereas others induce symptoms on N. glutinosa reminiscent of TAV (D. W. Mossop & R. I. B. Francki, unpublished data).
Alfalfa mosaic virus may also be confused with cucumber mosaic virus because it too is widespread, causes similar symptoms in some hosts and is transmitted by aphids in the non-persistent manner (Hull, 1969). However, it has small bacilliform particles, is antigenically distinct from cucumoviruses and invades C. amaranticolor and C. quinoa systemically (Bos & Jaspars, 1971).
Figures

Cucumber cv. Polaris systemically infected by the Q strain.

Symptoms on an infected field-grown plant of zucchini marrow.

Tomato cv. Rutgers systemically infected by an isolate from passionfruit.

N. glutinosa showing mild mosaic and leaf curling after infection by an isolate from spinach.

N. glutinosa showing chlorotic lesions on an inoculated leaf and systemic chlorosis after infection by the M strain.

N. glutinosa showing prominent mosaic symptoms after infection by an isolate from Capsicum sp.

Cowpea leaf showing brown lesions after inoculation with an isolate from lettuce.
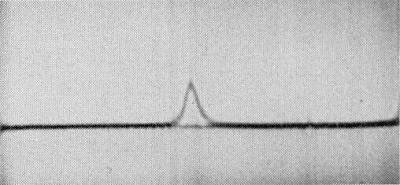
Schlieren pattern of the Q strain in an ultracentrifuge using a double-sector cell. (Sedimentation is from left to right.)

Separation of RNA components of the Q strain by polyacrylamide-gel electrophoresis under denaturing conditions (Pinder et al., 1974). (Left) viral RNA (Q strain), (right) marker RNA species, (centre) mixture of viral and marker RNA species. Markers were: TMV RNA (2.05 x 106 daltons; Boedtker, 1971), bacteriophage MS2 RNA (1.15 x 106 daltons; Fiers et al., 1976) and Escherichia coli ribosomal RNA (1.07 x 106 daltons (23 S), and 0.55 x 106 daltons (16 S); Stanley & Bock, 1965). Viral RNA-2 (Q2) was not resolved from MS2 RNA.

Purified preparation of an isolate from Capsicum sp. stained with uranyl acetate. Bar represents 100 nm. Inset shows an enlarged particle seen down its local two-fold axis, and rotated through 180°.

Part of a cell from a N. clevelandii leaf infected with the M strain showing electron-dense virus particles scattered in the cytoplasm. The densely staining cores of cytoplasmic ribosomes have been eliminated by treatment with ribonuclease (Hatta & Francki, 1979). Bar represents 500 nm.
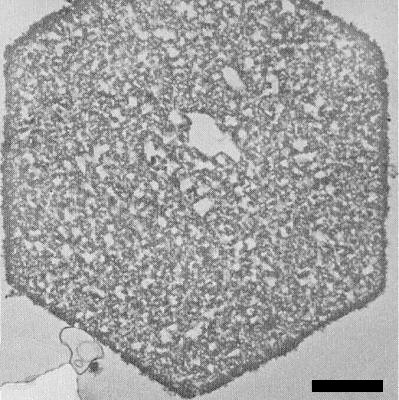
A virus crystal in the vacuole of a cell from a N. clevelandii leaf infected with an isolate from Capsicum sp. Bar represents 1 µm.
References list for DPV: Cucumber mosaic virus (213)
- Badami, Ann. appl. Biol. 46: 554, 1958.
- Bhargava, Ann. appl. Biol. 38: 377, 1951.
- Blencowe & Caldwell, Ann. appl. Biol. 36: 320, 1949.
- Boedtker, Biochim. biophys. Acta 240: 448, 1971.
- Bos & Jaspars, CMI/AAB Descriptions of Plant Viruses 46, 4 pp., 1971.
- Christie & Edwardson, Monograph Ser. Fla agric. Exp. Stn No. 9: 89, 1977.
- Clark, Peden & Symons, Virology 62: 434, 1974.
- Crowley, Aust. J. biol. Sci. 7: 141, 1954.
- Devergne, Meded. Faculteit Landbouwwetenschappen Rijks-Univ. Gent 40: 19, 1975.
- Devergne & Cardin, Annls Phytopath. 2: 639, 1970.
- Devergne & Cardin, Annls Phytopath. 5: 409, 1973.
- Devergne & Cardin, Annls Phytopath. 7: 255, 1975.
- Diaz-Ruiz & Kaper, Virology 80: 204, 1977.
- Doolittle, Phytopathology 6: 145, 1916.
- Dupont, Horn, Yot-Dauthy & Bové, C.r. Acad. Sci., Paris, D 267: 1013, 1968.
- Fiers, Contreras, Duerinck, Haegeman, Iserentant, Merregaert, Min Jou, Molemans, Raeymaekers, Van den Berghe, Volckaert & Ysebaert, Nature, Lond. 260: 500, 1976.
- Finch, Klug & Van Regenmortel, J. molec. Biol. 24: 303, 1967.
- Francki, Virology 24: 193, 1964.
- Francki, Virology 34: 694, 1968.
- Francki, in Principles and Techniques in Plant Virology, ed. C. I. Kado & H. Agrawal, New York: Van Nostrand-Reinhold, p. 295, 1972.
- Francki & Habili, Virology 48: 309, 1972.
- Francki, Randles, Chambers & Wilson, Virology 28: 729, 1966.
- Gerola, Bassi & Belli, Caryologia 18: 567, 1965.
- Gonda & Symons, Virology 88: 361, 1978.
- Gould & Symons, Nucleic Acids Res. 4: 3787, 1977.
- Gould & Symons, Eur. J. Biochem. 91: 269, 1978.
- Gould, Palukaitis, Symons & Mossop, Virology 84: 443, 1978.
- Govier, Ann. appl. Biol. 45: 62, 1957.
- Graham, Virology 3: 427, 1957.
- Habili & Francki, Virology 57: 392, 1974a.
- Habili & Francki, Virology 61: 443, 1974b.
- Habili & Francki, Virology 64: 421, 1975.
- Hatta & Francki, Virology 93: 265, 1979.
- Hoggan, J. agric. Res. 47: 689, 1933.
- Hollings, Ann. appl. Biol. 43: 86, 1955.
- Hollings & Stone, CMI/AAB Descriptions of Plant Viruses 79, 4 pp., 1971.
- Holmes, Virology 2: 611, 1956.
- Honda & Matsui, Phytopathology 58: 1230, 1968.
- Honda & Matsui, Phytopathology 64: 534, 1974.
- Hull, Adv. Virus Res. 15: 365, 1969.
- Jacrot, Chauvin & Witz, Nature, Lond. 266: 417, 1977.
- Jagger, Phytopathology 6: 148, 1916.
- Kaper, The Chemical Basis of Virus Structure, Dissociation and Reassembly (Frontiers of Biology Vol. 39), Amsterdam: North-Holland, 485 pp., 1975.
- Kaper & Diaz-Ruiz, Virology 80: 214, 1977.
- Kaper & Re, Virology 60: 308, 1974.
- Kaper & Waterworth, Science, N.Y. 196: 429, 1977.
- Kaper, Diener & Scott, Virology 27: 54, 1965.
- Kaper, Tousignant & Lot, Biochem. biophys. Res. Comm. 72: 1237, 1976.
- Kennedy, Day & Eastop, A Conspectus of Aphids as Vectors of Plant Viruses, London: Commonwealth Institute of Entomology, 114 pp., 1962.
- Kohl & Hall, J. gen. Virol. 25: 257, 1974.
- Komuro, Viruses in Vegetables. Tokyo: Seibundo Shinkosha, 300 pp., 1973.
- Linnasalmi & Toiviainen, Ann. Agric. Fenn. (Ser. Phytopath.) 13: 79, 1974.
- Lot & Kaper, Virology 74: 209, 1976.
- Lot, Jonard & Richards, FEBS Letters 80: 395, 1977.
- Lot, Marrou, Quiot & Esvan, Annls Phytopath. 4: 25, 1972.
- Lot, Marchoux, Marrou, Kaper, West, Van Vloten-Doting & Hull, J. gen. Virol. 22: 81, 1974.
- Lyons & Allen, J. Ultrastruct. Res. 27: 198, 1969.
- Matthews, Intervirology, 12: 132, 1979.
- Mink, CMI/AAB Descriptions of Plant Viruses 92, 4 pp., 1972.
- Mossop & Francki, Virology 81: 177, 1977.
- Mossop & Francki, Virology 86: 562, 1978.
- Mossop & Francki, Virology 94: 243, 1979.
- Mossop, Francki & Grivell, Virology 74: 544, 1976.
- Neergaard, Seed Pathology, Vol. I, London: Macmillan, 839 pp., 1977.
- Normand & Pirone, Virology 36: 538, 1968.
- Peden & Symons, Virology 53: 487, 1973.
- Phatak, Diaz-Ruiz & Hull, Phytopath. Z. 87: 132, 1976.
- Pinder, Staynov & Gratzer, Biochemistry 13: 5373, 1974.
- Price, Phytopathology 24: 743, 1934.
- Price, Am. J. Bot. 27: 530, 1940.
- Richards, Jonard, Jacquemond & Lot, Virology 89: 395, 1978.
- Schmelzer, Phytopath. Z. 28: 1, 1957.
- Schmelzer, CMI/AAB Descriptions of Plant Viruses 65, 3 pp., 1971.
- Schwinghamer & Symons, Virology 79: 88, 1977.
- Scott, Virology 20: 103, 1963.
- Scott, Virology 34: 79, 1968.
- Simons, Phytopathology 45: 217, 1955.
- Simons, Phytopathology 47: 145, 1957.
- Simons, Virology 9: 612, 1959.
- Smith, A Textbook of Plant Virus Diseases (3rd Ed.), London: Longman, 684 pp., 1972.
- Stanley & Bock, Biochemistry 4: 1302, 1965.
- Symons, Mol. Biol. Reptr. 2: 277, 1975.
- Takanami, Kubo & Imaizumi, Virology 80: 376, 1977.
- Tolin, in The Atlas of Insect and Plant Viruses, ed. K. Maramorosch, New York: Academic Press, p. 303, 1977.
- Tomlinson & Carter, Ann. appl. Biol. 66: 381, 1970.
- Van Regenmortel, Virology 31: 391, 1967.
- Van Regenmortel, Hendry & Baltz, Virology 49: 647, 1972.
- Watson & Roberts, Proc. R. Soc. Ser. B. 127: 543, 1939.